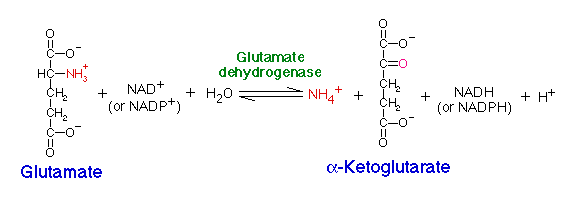
Glutamate dehydrogenase (GDH) is located in the mitochondria and is an important branch-point enzyme between carbon and nitrogen metabolism (13). GDH catalyzes the reversible NAD (P)+-linked oxidative deamination of L-glutamate into alpha ketoglutarate and ammonia in two steps. The first step involves a Schiff base intermediate being formed between ammonia and alpha ketoglutarate. This Schiff base intermediate is crucial because it establishes the alpha carbon atom in glutamate’s stereochemistry. The second step involves the Schiff base intermediate being protonated, which is done by the transfer of a hydride ion from NADPH resulting in L-glutamate. GDH is unique because it is able to utilize both NAD+ and NADP+ (15). NADP+ is utilized in the forward reaction of alpha ketogluterate and free ammonia, which are converted to L-glutamate via a hydride transfer from NADPH to glutamate (15). NAD+ is utilized in the reverse reaction, which involves L-glutamate being converted to alpha ketoglutarate and free ammonia via an oxidative deamination reaction (1). The extensive production of ammonia by peripheral tissue or glutamate dehydrogenase is not allowed because of the highly toxic effects of circulating ammonia in cells. Therefroe, the ammonia produced in the reverse reaction of GDH is excreted as NH4+ in the urine, by first going through the urea cycle.
Figure 1: Reversible reaction catalyzed by GDH
GDH is a hexamer of 449 residues with a molecular weight of 49.2 kD (18). All six subunits are identical in GDH, and contain two distinct domains, that are roughly the same size, and are separated by a large active site cleft (9). The overall structure of GDH is composed of eighteen alpha helices and thirteen beta sheets, which are both parallel and anti-parallel. As common to most oxidoreductases the beta sheets are flanked by a layer of alpha helices (18). The free energy change for the conversion of glutamate to alpha ketoglutarate is 3.7 kcal/mol (4). Although this ATP-consuming pathway is energetically unfavourable, this could be related to the continuation of re-dox equilibrium to re-oxidise the excess of NADH produced during glycolysis.
JMol Image of Glutamate Dehydrogenase (1V9L)
| The secondary structure of GDH is identical in all six subunits. Each subunit contains eighteen alpha helices, and thirteen beta sheets, which are both parallel and anti-parallel. |
GDH is allosterically regulated by the cell’s energy state. During the formation of alpha ketoglutarate GDP and ADP positively regulate GDH in mammals, and GTP, ATP, leucine, and coenzyme inhibit the enzyme. Therefore, when the level of ATP is high, conversion of glutamate to alpha ketoglutarate is limited; however when the cellular energy charge is low, glutamate is converted to ammonia and alpha ketoglutarate (2).
GDH consist of two domains, which are separated by an active site cleft, which also contains the substrate-binding site at the depth of the cleft (9). Domain I is made up of residues 4-181 and 400-421, and is responsible for directing the assembly of the subunits into a hexamer. Domain II makes up the glutamate-binding domain, and is composed of mainly beta sheets involving residues 182-399 (5). Domain I is also called the C-domain, whereas the Domain II is called the N-domain (9). Each domain is constructed from a central beta sheet that is flanked by alpha helices. The C-domain is composed of an alpha/beta super-secondary structure, which is common of the Rossmann fold. Whereas the N-domain is composed of an alpha/beta super-secondary structure, however is not like the Rossman fold. The NAD+ cofactor binds at the C-terminal end of the parallel beta strands in the N-domain, lying in the cleft between the N and C-domains. The glutamate substrate binds deep in this cleft, with the side chain of the glutamate lying in a pocket on the enzyme surface. Residues 193-204, and 383-393 are essential for the glutamate to bind in the cleft (1).
NAD+ binding domains are located on top of the N and C-domains. These NAD+ binding domains rotate down upon the substrate and coenzyme to initiate catalysis. The NAD+ binding domain contains a forty eight-residue “antenna” that extends from the top of the NAD+ binding domain. This antenna undergoes conformational changes as the cleft of the active site opens and closes (2). Both of the domains are positioned differently in GDH. When GDH is not bound by glutamate its cleft is open, however when GDH is bound by glutamate it is closed. This position difference between the two domains allows the cleft to be closed, which brings the C4 of the nicotinamide ring and the alpha carbon of the glutamate substrate into the appropriate orientation for a hydride transfer to occur. Residues 200-206, 375-379, and 421-423 are critical for the control of the hinges that open or close the cleft between the two domains (1). The residues that form this hinge, which allow the cleft to open or close are both near and far from the active site. Gly 182, 183, Leu 185, and Gly 186 are near the active site, whereas Gly 361 is far from the active site. These five residues form two long loops connecting the two domains (9).
The first step in the mechanism for catalytic activity of GDH is the -deprotonation of the alpha-amino group of glutamate by Asp 165, which acts as a general base. Next, a hydride transfer to NAD+ occurs, which forms a Schiff base intermediate (13). During the first step a large movement between C-domain and N-domain occurs, which closes the cleft and brings C4 of the nicotinamide ring and the alpha carbon of the substrate into the correct position for a hydride transfer (1). The second step involves the attack of a water molecule on the Schiff base intermediate, which is enhanced by Lys 125. The direction of the attack is very specific, so that the stereochemistry of the developing carbinolamine will be the L isomer and not the D isomer. During the generation of the carbinolamine intermediate and its conversion to alpha ketoglutarate, residue Asp 165 is very crucial for the transfer of the protons to and from the substrate. The final step that GDH catalyzes involves the loss of a single proton from each Lys 125 and Asp 165, which is transferred from water to GDH (13).